Strictly speaking, species diversity is the number of different species in a particular area (species richness) weighted by some measure of abundance such as number of individuals or biomass. However, it is common for conservation biologists to speak of species diversity even when they are actually referring to species richness.
Another measure of species diversity is the species evenness, which is the relative abundance with which each species is represented in an area. An ecosystem where all the species are represented by the same number of individuals has high species evenness. An ecosystem where some species are represented by many individuals, and other species are represented by very few individuals has a low species evenness. Table 6.1 shows the abundance of species (number of individuals per hectare) in three ecosystems and gives the measures of species richness (S), evenness (E), and the Shannon diversity index (H).
Shannon's diversity index H=–(∑(ρilnρi))
ρi
is the proportion of the total number of specimens
i expressed as a proportion of
the total number of species for all species in the
ecosystem. The product of
ρilnρi
for each species in the ecosystem is summed, and multiplied
by
–1
to give H. The species
evenness index
( E)
is calculated as
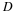 .
.
Hmax
is the maximum possible value of
H, and is
equivalent to
lnS.
Thus
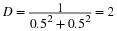
See Gibbs et al., 1998: p157 [link] and Beals et al. (2000) [link] for discussion and examples. Magurran (1988) [link] also gives discussion of the methods of quantifying diversity.
In Table 6.1, ecosystem A shows the greatest diversity in terms of species richness. However, ecosystem B could be described as being richer insofar as most species present are more evenly represented by numbers of individuals; thus the species evenness (E) value is larger. This example also illustrates a condition that is often seen in tropical ecosystems, where disturbance of the ecosystem causes uncommon species to become even less common, and common species to become even more common. Disturbance of ecosystem B may produce ecosystem C, where the uncommon species 3 has become less common, and the relatively common species 1 has become more common. There may even be an increase in the number of species in some disturbed ecosystems but, as noted above, this may occur with a concomitant reduction in the abundance of individuals or local extinction of the rarer species.
Species richness and species evenness are probably the most frequently used measures of the total biodiversity of a region. Species diversity is also described in terms of the phylogenetic diversity, or evolutionary relatedness, of the species present in an area. For example, some areas may be rich in closely related taxa, having evolved from a common ancestor that was also found in that same area, whereas other areas may have an array of less closely related species descended from different ancestors (see further comments in the section on Species diversity as a surrogate for global biodiversity).
To count the number of species, we must define what constitutes a species. There are several competing theories, or "species concepts" (Mayden, 1997 [link]). The most widely accepted are the morphological species concept, the biological species concept, and the phylogenetic species concept.
Although the morphological species concept (MSC) is largely outdated as a theoretical definition, it is still widely used. According to this concept:
species are the smallest groups that are consistently and persistently distinct, and distinguishable by ordinary means. (Cronquist, 1978 [link]).
In other words, morphological species concept states that "a species is a community, or a number of related communities, whose distinctive morphological characters are, in the opinion of a competent systematist, sufficiently definite to entitle it, or them, to a specific name" (Regan, 1926: 75 [link]).
The biological species concept (BSC), as described by Mayr and Ashlock (1991) [link], states that
"a species is a group of interbreeding natural populations that is reproductively isolated from other such groups".
According to the phylogenetic species concept (PSC), as defined by Cracraft (1983) [link], a species :
"is the smallest diagnosable cluster of individual organism [that is, the cluster of organisms are identifiably distinct from other clusters] within which there is a parental pattern of ancestry and descent".
These concepts are not congruent, and considerable debate exists about the advantages and disadvantages of all existing species concepts (for further discussion, see the module on Macroevolution: essentials of systematics and taxonomy).
In practice, systematists usually group specimens together according to shared features (genetic, morphological, physiological). When two or more groups show different sets of shared characters, and the shared characters for each group allow all the members of that group to be distinguished relatively easily and consistently from the members of another group, then the groups are considered different species. This approach relies on the objectivity of the phylogenetic species concept (i.e., the use of intrinsic, shared, characters to define or diagnose a species) and applies it to the practicality of the morphological species concept, in terms of sorting specimens into groups (Kottelat, 1995 [link], 1997 [link]).
Despite their differences, all species concepts are based on the understanding that there are parameters that make a species a discrete and identifiable evolutionary entity. If populations of a species become isolated, either through differences in their distribution (i.e., geographic isolation) or through differences in their reproductive biology (i.e., reproductive isolation), they can diverge, ultimately resulting in speciation. During this process, we expect to see distinct populations representing incipient species - species in the process of formation. Some researchers may describe these as subspecies or some other sub-category, according to the species concept used by these researchers. However, it is very difficult to decide when a population is sufficiently different from other populations to merit its ranking as a subspecies. For these reasons, subspecific and infrasubspecific ranks may become extremely subjective decisions of the degree of distinctiveness between groups of organisms (Kottelat, 1997 [link]).
An evolutionary significant unit (ESU) is defined, in conservation biology, as a group of organisms that has undergone significant genetic divergence from other groups of the same species. According to Ryder, 1986 [link] identification of ESUs requires the use of natural history information, range and distribution data, and results from analyses of morphometrics, cytogenetics, allozymes and nuclear and mitochondrial DNA. In practice, many ESUs are based on only a subset of these data sources. Nevertheless, it is necessary to compare data from different sources (e.g., analyses of distribution, morphometrics, and DNA) when establishing the status of ESUs. If the ESUs are based on populations that are sympatric or parapatric then it is particularly important to give evidence of significant genetic distance between those populations.
ESUs are important for conservation management because they can be used to identify discrete components of the evolutionary legacy of a species that warrant conservation action. Nevertheless, in evolutionary terms and hence in many systematic studies, species are recognized as the minimum identifiable unit of biodiversity above the level of a single organism (Kottelat, 1997 [link]). Thus there is generally more systematic information available for species diversity than for subspecific categories and for ESUs. Consequently, estimates of species diversity are used more frequently as the standard measure of overall biodiversity of a region.
Global biodiversity is frequently expressed as the total number of species currently living on Earth, i.e., its species richness. Between about 1.5 and 1.75 million species have been discovered and scientifically described thus far (LeCointre and Guyader, 2001 [link]; Cracraft, 2002 [link]). Estimates for the number of scientifically valid species vary partly because of differing opinions on the definition of a species.For example, the phylogenetic species concept recognizes more species than the biological species concept. Also, some scientific descriptions of species appear in old, obscure, or poorly circulated publications. In these cases, scientists may accidentally overlook certain species when preparing inventories of biota, causing them to describe and name an already known species.
More significantly, some species are very difficult to identify. For example, taxonomically "cryptic species" look very similar to other species and may be misidentified (and hence overlooked as being a different species). Thus, several different, but similar-looking species, identified as a single species by one scientist, are identified as completely different species by another scientist. For further discussion of cryptic species, with specific examples of cryptic frogs from Vietnam, see Inger (1999) [link] and Bain et al., (in press) [link].
Scientists expect that the scientifically described species represent only a small fraction of the total number of species on Earth today. Many additional species have yet to be discovered, or are known to scientists but have not been formally described. Scientists estimate that the total number of species on Earth could range from about 3.6 million up to 117.7 million, with 13 to 20 million being the most frequently cited range (Hammond, 1995 [link]; Cracraft, 2002 [link]).
The estimation of total number of species is based on extrapolations from what we already know about certain groups of species. For example, we can extrapolate using the ratio of scientifically described species to undescribed species of a particular group of organisms collected from a prescribed area. However, we know so little about some groups of organisms, such as bacteria and some types of fungi, that we do not have suitable baseline data from which we can extrapolate our estimated total number of species on Earth. Additionally, some groups of organisms have not been comprehensively collected from areas where their species richness is likely to be richest (for example, insects in tropical rainforests). These factors, and the fact that different people have used different techniques and data sets to extrapolate the total number of species, explain the large range between the lower and upper figures of 3.6 million and 117.7 million, respectively.
While it is important to know the total number of species of Earth, it is also informative to have some measure of the proportional representation of different groups of related species (e.g. bacteria, flowering plants, insects, birds, mammals). This is usually referred to as the taxonomic or phylogenetic diversity. Species are grouped together according to shared characteristics (genetic, anatomical, biochemical, physiological, or behavioral) and this gives us a classification of the species based on their phylogenetic, or apparent evolutionary relationships. We can then use this information to assess the proportion of related species among the total number of species on Earth. Table 6.1 contains a selection of well-known taxa.
Most public attention is focused on the biology and ecology of large, charismatic species such as mammals, birds, and certain species of trees (e.g., mahogany, sequoia). However, the greater part of Earth's species diversity is found in other, generally overlooked groups, such as mollusks, insects, and groups of flowering plants.
Mayden, R.L. (1997). A hierarchy of species concepts: the denouement in the saga of the species problem. In M.F. Claridge, H.A. Dawah, and M.R. Wilson (Eds.), Species: the units of biodiversity. (pp. 381-424). London, U.K.: Chapman and Hall.
Du Rietz, G.E. (1930). The fundamental units of biological taxonomy. Svensk Botanisk Tidskrift, 24, 333-428.
Mayr, E. (1982). The growth of biological thought: diversity, evolution, and inheritance. Cambridge, Massachusetts, U.S.A.: Harvard University Press.
Bisby, F.A. and J. Coddington. (1995). Biodiversity from a taxonomic and evolutionary perspective. In V.H. Heywood and R.T. Watson (Eds.), Global Biodiversity Assessment. (pp. 27-56). Cambridge, U.K: Cambridge University Press.
Cracraft, C. (1983). Species concepts and speciation analysis. Ornithology, 1, 159-187.
Kottelat, M. (1995). Systematic studies and biodiversity: the need for a pragmatic approach. Journal of Natural History, 29, 565-569.
Kottelat, M. (1997). European freshwater fishes: an heuristic checklist of the freshwater fishes of Europe (exclusive of former USSR), with an introduction for non-systematists and comments on nomenclature and conservation. Biologia (Bratislava), 52 (Supplement 5), 1-271.
Ryder, O.A. (1986). Species conservation and systematics: the dilemma of subspecies. Trends in Ecology and Evolution, 1(1), 9-10.
Cracraft, C. (2002). The seven great questions of systematic biology: an essential foundation for conservation and the sustainable use of biodiversity. Annals of the Missouri Botanical Garden, 89, 127-144.
Lecointre, G. and H. Le Guyader. (2001). Classification phylogenetique du vivant. Paris, France: Belin.
Inger, R.F. (1999). Distribution of amphibians of southern Asia and adjacent islands. (pp. 445-482). Baltimore, Maryland, U.S.A.: Johns Hopkins University Press.
Bain, R.H., A. Lathrop, R.W. Murphy, N.L. Orlov, and Ho Thu Cuc. (in press). Cryptic species of a cascade frog from Southeast Asia: taxonomic revisions and descriptions of six new species. American Museum Novitates.
Hammond, P. (1995). The current magnitude of biodiversity. In V.H. Heywood and R.T. Watson (Eds.), Global Biodiversity Assessment. (pp. 113-138). Cambridge, U.K: Cambridge University Press.
Beals, M., L. Gross, S. Harrell. (1998). Diversity indices: Shannon's H and E. In L.J. Gross, B.C. Mullin, S.E. Riechert, O.J. Schwarz, M. Beals, S. Harrell (Ed.), Alternative routes to quantitative literacy for the life sciences - A project supported by the National Science Foundation. Knoxville, Tennessee: University of Tennessee, Knoxville.
Gibbs, J.P., M.L. Hunter, Jr. and E.J. Sterling. (1998). Problem-Solving in conservation biology and wildlife management. In Exercises for class, field and laboratory. Blackwell Science, Massachusetts, U.S.A
Magurran, A.E. (1998). Ecological diversity and its measurement. In Princeton, New Jersey, U.S.A: Princeton University Press.
Cronquist, A. (1978). Once again, what is a species? In L.V. Knutson (Ed.), Biosystematics in Agriculture. (pp. 3-20). Montclair, New Jersey, U.S.A: Allenheld Osmin.
Regan, C.T. (1926). Organic Evolution. In Report British Association for Advancement of Science, 1925. (pp. 75-86).
Mayr, E. and P.D. Ashlock. (1991). Principles of systematic zoology. New York, New York, U.S.A.: McGraw-Hill.
a species is a group of interbreeding natural populations unable to successfully mate or reproduce with other such groups, and which occupies a specific niche in nature (Mayr, 1982; Bisby and Coddington, 1995).
Ecosystema community plus the physical environment that it occupies at a given time.
Evolutionary significant unita group of organisms that has undergone significant genetic divergence from other groups of the same species. Identification of ESUs is based on natural history information, range and distribution data, and results from analyses of morphometrics, cytogenetics, allozymes and nuclear and mitochondrial DNA. Concordance of those data, and the indication of significant genetic distance between sympatric groups of organisms, are critical for establishing an ESU.
Morphological species conceptspecies are the smallest natural populations permanently separated from each other by a distinct discontinuity in the series of biotype (Du Rietz, 1930; Bisby and Coddington, 1995).
Parapatricoccupying contiguous but not overlapping ranges.
Phylogenetic diversitythe evolutionary relatedness of the species present in an area.
Phylogenetic species concepta species is the smallest group of organisms that is diagnosably [that is, identifiably] distinct from other such clusters and within which there is a parental pattern of ancestry and descent (Cracraft, 1983; Bisby and Coddington, 1995).
Species diversitythe number of different species in a particular area (i.e., species richness) weighted by some measure of abundance such as number of individuals or biomass.
Species evennessthe relative abundance with which each species are represented in an area.
Species richnessthe number of different species in a particular area
Sympatricoccupying the same geographic area.